過去の研究テーマ
「脊椎動物の進化史を遺伝子で解明する」
ダーウィンは進化論の中で、種は分岐を繰り返しながら多様化を遂げてきたことを説いていますが、その分岐の歴史を1つの樹のように描いたものが「系統樹」です。この系統樹によって、地球上の生物がどのような順序で分岐を繰り返してきたのか、どの生物どうしがお互いに近縁なのかを理解することが可能になります。つまり系統樹は、進化学の中でもっとも基本的かつ重要な情報を我々に提示してくれます。かつては、生物の「かたち」を指標とした系統推定も、現在ではより客観的な「DNA配列」によるものが主流になりつつあります(これを分子系統学と呼びます)。分子系統学 が隆盛となった頃(2000年前後)は、かたちに基づく伝統的な系統樹と、DNAに基づく系統樹の間に矛盾が生じることが多く、しばしば論争が繰り広げられてきました。これらの論争の多くは、生物の系統樹に関わる最終結論を導く上で大きな役割を果たしましたが、それ以上に生物の「かたち」が種の分岐に伴ってどのように進化しうるのか、そのメカニズムを明らかにするための新たなる学問分野(エボデボ)を創出するという大きな役割を果たしました。下記に紹介する系統推定に関わるこれまでの研究も、まさに私たちが今進めているエボデボ研究をおこなう上で重要な基盤となったものです。まだまだ、系統関係が明らかになっていない生物種も少なくありません。今後も系統推定に関わる研究は進化学の中でも重要な役割を果たすことは間違いありません。
クジラの起源
クジラ類は哺乳類であるため、その祖先は陸上を四肢で闊歩し、また、他の陸上哺乳類と同じように全身が体毛で包まれていたはずです。しかし現在のクジラ類に四肢はなく、前肢は遊泳をするための鰭に変化し、後肢は完全に退化しています。さらに重力から解放された体は巨大化を続け、水中での抵抗を限りなく減らすように体毛は消失し、美しい流線型をなしています。このクジラはいったいどんな動物から派生してきたのでしょうか?
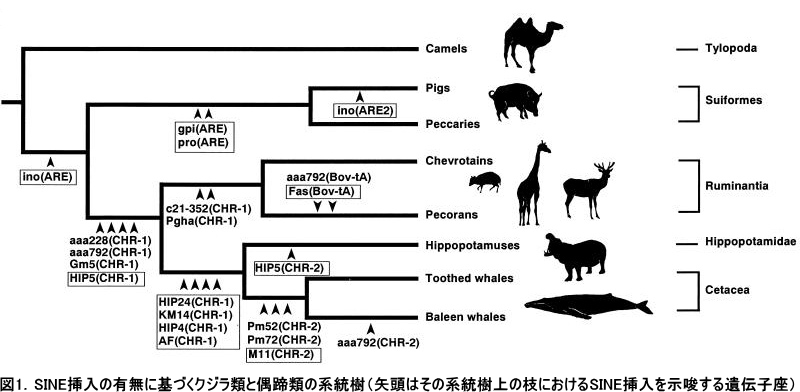
私の所属していた研究室(岡田典弘教授)では、レトロポゾンの1種であるSINEの挿入を指標として、鯨類の分子系統樹の構築を試みました。そしてSINE法による解析では、クジラ類と偶蹄類の共通祖先の中で、もっとも最初に分岐したのは核脚類(ラクダやラマの仲間)であり、その次にブタやペッカリーのグループが分岐し、続いて反芻類(ウシ、シカ、キリンの仲間)が分岐したことがわかりました。つまりクジラ類とカバ類はその後も共通祖先を共有していたことになり、最終的にクジラ類にもっとも近縁な現生哺乳類はカバであることが明らかとなりました(Nikaido, Rooney and Okada 1999 PNAS; 鯨偶蹄類の系統樹に関しては図1を参照)。
我々の提言した「カバとクジラ類の近縁性」によってもたらされた最も大きな問題は、化石記録との不整合性でした。分子データが示唆するように、クジラ類がカバと姉妹群を形成すると仮定した場合、形態学的にはクジラ類に近縁だと考えられているメソニクス類も当然、偶蹄目の内部に入り込むことになります。そうすると、ここである大きな疑問が生じてきます。それは、メソニクス類の化石記録はおよそ6000万年以上も前の地層から数多く発見されているにもかかわらず、原始的な偶蹄類の化石は5400万年ほど、加えてクジラ類の化石も同様に5000万年程度しかさかのぼらないことです。もしメソニクス類がクジラ類とともに偶蹄類の内部に入り込むとしたら、初期の偶蹄類の化石記録において大きな空白が生じてしまうことになります。つまり、古生物学的な定説とも言われてきたメソニクス類とムカシクジラ類との近縁性が揺らぎ始めたのです。
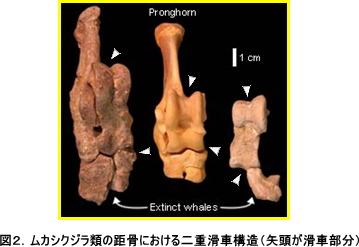
そして、古生物学的な方面からもメソニクスやクジラ類(ムカシクジラ類)における形態形質を再検討する動きも見られ始め、さらには形態学の分野では伝統的な仮説であるメソニクスとクジラ類の近縁性にも疑問を投げかけるような発見がありました。そこで非常に重要なキーポイントとなるのが、偶蹄目を定義付ける上で最も重要な形質の1つである距骨の二重滑車構造です。つまり、偶蹄類に存在する距骨の二重滑車構造がメソニクスに存在しないことが判っているので、それでは逆にその構造がクジラ類に存在するのであれば、偶蹄類とクジラ類がより近縁である強い証拠になると同時に、逆にメソニクスとクジラ類の近縁性は薄らいでいくはず、というわけです。ムカシクジラ類における距骨の発見は2チームが独立に報告したもので、それぞれが2001年9月付けのNatureとScience誌に掲載されました。彼らはムカシクジラ類の全身骨格の復元に成功し、その距骨に明らかな二重滑車構造をみいだしました(図2参照)。さらには今回発掘したムカシクジラ類の1種の後肢の構造が、カバに近縁な絶滅種であるアントラコテリウム類のそれと非常に似通っていることを挙げ、今後のさらなる古生物学的な発見が続けば、カバとクジラ類の近縁性が形態の分野からも支持される日が来るのかもしれないと主張しました。これまで、鯨類と偶蹄類は別々な目に分類されてきましたが、我々を含めた鯨類の系統に関する新発見に基づき、あらたなる分類群「鯨偶蹄目」が提唱され、現在の図鑑などではほぼこの分類に統一されています。
人類史の始まりと共に続いてきた、いわば哺乳類進化の命題でもあるクジラの起源に関する謎は、分子系統学と古生物学の見事な調和により、ようやく解き明かされつつあるのではないでしょうか。
ハクジラ論争
1990年代は、いわば分子系統学が隆盛を極めた時代ともいえます。その頃は、古生物学や形態学などで提唱されてきたあらゆる系統仮説が次々とDNAデータによって刷新されていき、DNAのデータに基づく系統樹は往々にして正しい、という分子系統学者達の若干の過信もあった時代にも思えます。そのような状況の中で、分子系統学サイドの弱点を露呈してしまうような論争がありました。それがハクジラ類の単系統論争です。
現生のクジラ類は形態学的に、歯をもつハクジラ類と髭板をもつヒゲクジラ類に大きく2つのグループに分けられてきました。歯や髭板の有無以外にも、テレスコーピング様式の違いに起因する頭がい骨の違い、左右の非対称性、メロン体の有無など、様々な特徴がハクジラ類とヒゲクジラ類それぞれの単系統性を強く示唆するものでした。しかし、1993年にMilinkovitchらはミトコンドリアのcytb遺伝子の部分配列を用いた分子系統解析により、ハクジラ類の中でも特に大型のマッコウクジラが他のハクジラよりもむしろヒゲクジラ類に近縁である、という驚くべき仮説をNatureに発表しました。分子系統学隆盛の時代ではありましたが、このあまりにもチャレンジングな仮説に対しては形態学者らが一斉に反発し、大きなセンセーションとなりました。
Milinkovitchらはその後も、ハクジラ類の単系統性を示唆するいくつもの形態学的特徴が、実は解釈を変えればマッコウクジラとヒゲクジラ類の近縁性を示唆するものになりうると主張するなど、お互いの間では長い期間に渡っての論戦が続きました。ただ、Milinkovitchらの研究では、解析に用いる外群によって結果が変わってしまうことなどDNA配列の統計的処理による誤差が生じている可能性も指摘され、第三者的な手法による解析が待たれていました。
そこで我々は、DNAデータを利用しているものの、統計的な解析を一切必要としないSINE法(全項目を参照)を用いて、鯨類の系統樹を構築することを試みました。その結果、マッコウクジラが他のハクジラ類と単系統群を形成する遺伝子座が複数単離され、それに矛盾する遺伝子座は観察されませんでした(Nikaido et al. 2001 PNAS; 系統樹は図1を参照)。つまりSINE法による結果はDNAを用いた解析であるにも関わらず形態学的な伝統仮説を強く示唆するもので、この研究によって一連の論争に終止符が打たれました。ハクジラ類の単系統性に関する論争では、種の分岐にかかる時間が短かったり、もしくはその後の進化速度が一定でない場合には、十分なデータ量と適切な塩基置換モデルを用いて解析しないと、時として誤った系統樹を導き出してしまうこともありうるという教訓になりました。
我々のSINE法による解析ではさらに、これまでは1つのグループにまとまると考えられてきたカワイルカ類が実は多系統であり、特にガンジスカワイルカ類はアカボウクジラ類が分岐するより前に派生していた古い系統であることが判明しました(図1参照)。つまり、世界の河川に生息するカワイルカ類は見た目こそ似通っていますが(吻が長い、目が退化傾向にある、首が可動する等)、その形態的な類似性は河川という環境に適応することで平行的に獲得してきたものだということが明らかになったのです。
ヒゲクジラ系統樹
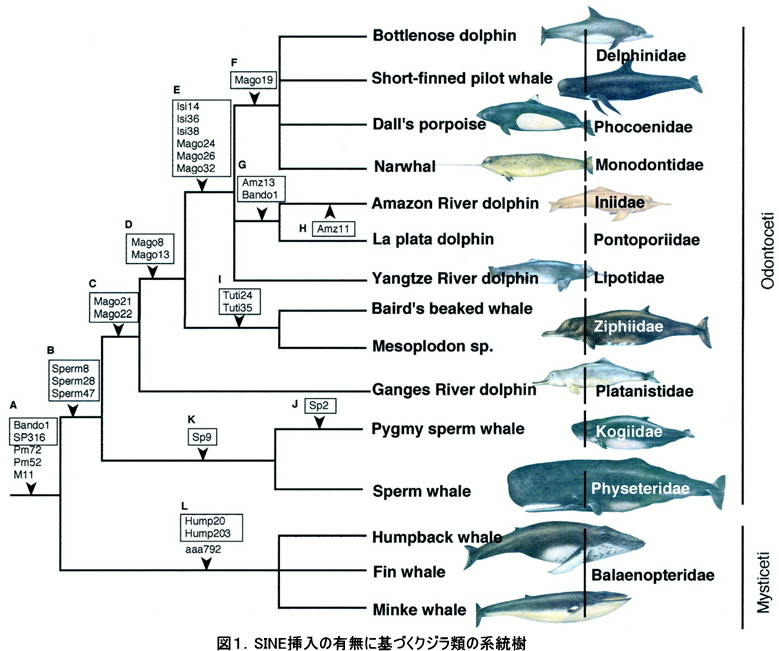
現生のヒゲクジラ亜目は、一般に大型な鯨類12種から構成されており、地球上最大の哺乳動物「シロナガスクジラ」を包含するグループです。ヒゲクジラ類は食用資源としてもその価値が大きく、古来より我々が慣れ親しんできた哺乳類のひとつでもあります。しかしヒゲクジラ類に関しては、形態・分子を含め様々な側面から系統学的研究がなされているにもかかわらず、それらの系統進化に関する問題の多くは解決に至っていませんでした。そこで私達は、レトロポゾンの一種であるSINEの挿入の有無を指標にして、ヒゲクジラ類全12種の系統解析をおこないました。我々の研究によって明らかになった結果は以下に要約されます。1.セミクジラとホッキョククジラが単系統で、現生ヒゲクジラ類の中で最初に分岐したグループである。2.コセミクジラ類はセミクジラ類と単系統群を形成せず、セミクジラ科の次に分岐したグループである。3.ザトウクジラとナガスクジラが単系統群を形成する。4.イワシクジラ、ニタリクジラが単系統でその姉妹群にシロナガスクジラが位置する(Nikaido et al. 2006 MBE; 系統樹は図1を参照してください)。以上の結果は我々が同時に進めてきたミトコンドリア全長配列を指標としたヒゲクジラ類の系統解析においても、まったく同じ樹形が得られたことから、この系統樹の信頼性が高いと考えられます。
ただ以上の結果に加えて、セミクジラ類が分岐した後の系統樹上において矛盾するSINE挿入パターンが検出されました。これは祖先多型現象によるものである事が予想されます。つまり、コセミクジラ類の分岐後に急激な放散が起きたことが示唆されます。ミトコンドリア全長配列を用いた解析では、上述の急速な放散が起きた時期は今からおよそ2000万年前と推定されました。現在までのところ、2000万年前の地球環境において、劇的な海流の変化や、海水面の変遷など特徴的な史実は明らかになっていませんが、地球規模の環境変化に伴うプランクトンの大量発生や、それを効率良く濾し取ることのできる様なひげ板の進化が相乗効果となって、放散が起きたのではないかと考えています。
コウモリの起源
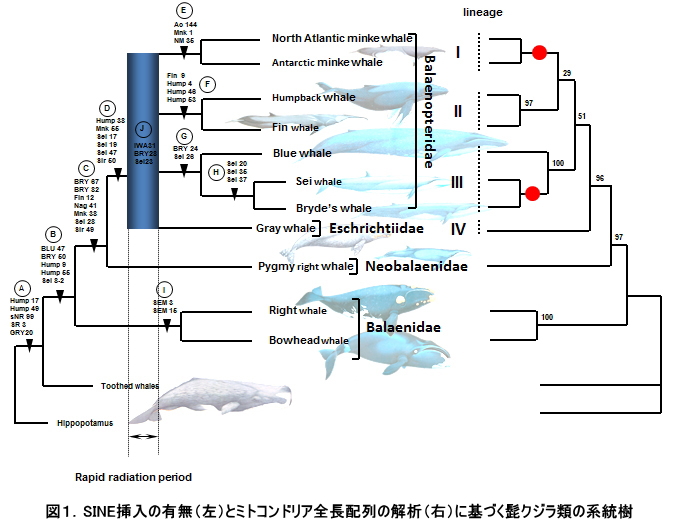
クジラ類と同等か、もしくはそれ以上に劇的な形態的特殊化を遂げた哺乳類としてコウモリ類(翼手目)が挙げられます。コウモリ類は前肢を翼へと変化させ、その一部の種は体重を哺乳類最少に匹敵するまでに軽量化させることで完全なる空中飛翔能力を獲得しました。一般に、このように劇的な形態進化を遂げた種の系統推定は極めて難しいのが常ですが、さらにコウモリ類の分類を難しくさせたのが、進化的な中間段階を示す化石記録が存在しないことでした。つまり最古のコウモリ類の化石として記載されているIcaronycteris indexはすでに完全なコウモリ様の形態を獲得しており、コウモリ類とその他の哺乳類をつなぐようなグループは見つかっていないため、コウモリ類がどの哺乳類と近縁なのかを推定するのは困難を極めたのです。
コウモリ類は、形態学的には霊長類、皮翼類(ヒヨケザル)そしてツパイと共にアーコンタと呼ばれる分類群にまとめられており、特に滑空はするが飛翔はしないヒヨケザルと近縁である可能性が高いと考えられてきました。また、コウモリ類の単系統性については疑う余地のないものとして扱われてきました。しかし、Pettigrewらは1986年にScienceに発表した論文において、コウモリ類の視神経のつながり方に注目し、大コウモリ類は小コウモリ類よりもむしろ霊長類に近縁である可能性を示唆し「空飛ぶ霊長類仮説」を提唱しました。つまりコウモリ類は単系統群を形成せず、飛翔能力や翼の獲得が哺乳類進化の過程で独立に2度起きたと主張したのです。このあまりにも突飛な仮説に対しては多くの反論がScience誌上に掲載され、大々的な論争が繰り広げられました。その結果、多くの研究者の間ではコウモリ類は単一起源であるという考えが主流になりつつありましたが、2000年当時の段階では十分な量のDNA配列とアウトグループを用いた系統解析は実施されておらず、さらにはそのコウモリ類自体はどの哺乳類に近縁なのかという問題も含めた包括的な結論は出されていませんでした。
そこで我々は、哺乳類の目間レベルでの包括的な系統解析に適したミトコンドリア全長配列を、大コウモリについて決定し、それらを他哺乳類のものと合わせて最尤法を用いて解析をおこないました(図1: Nikaido et al. 2000 JME )。その結果、大コウモリ類と小コウモリ類の単系統性は高い信頼度によって支持されました。つまりPettigrewらの主張する「空飛ぶ霊長類仮説」は、十分なデータ量を用いた我々の解析により完全に否定されたことになります。また、さらに重要なこととしてコウモリ類は有蹄類(ウシやイヌ、ウマなどのグループ)に近縁であり、伝統的にその近縁性が支持されてきた霊長類やヒヨケザルとは近縁ではないことが十分な信頼性を持って示されたのです。つまり、ヒヨケザルとコウモリ類は近縁であると考えられてきましたが、それらは滑空と飛翔によってもたらされた表層的な類似性によるものである可能性が高く(図2)、ここであらためてコウモリ類の形態に基づく系統推定の難しさが明らかとなったわけです。
タンザニアシーラカンス
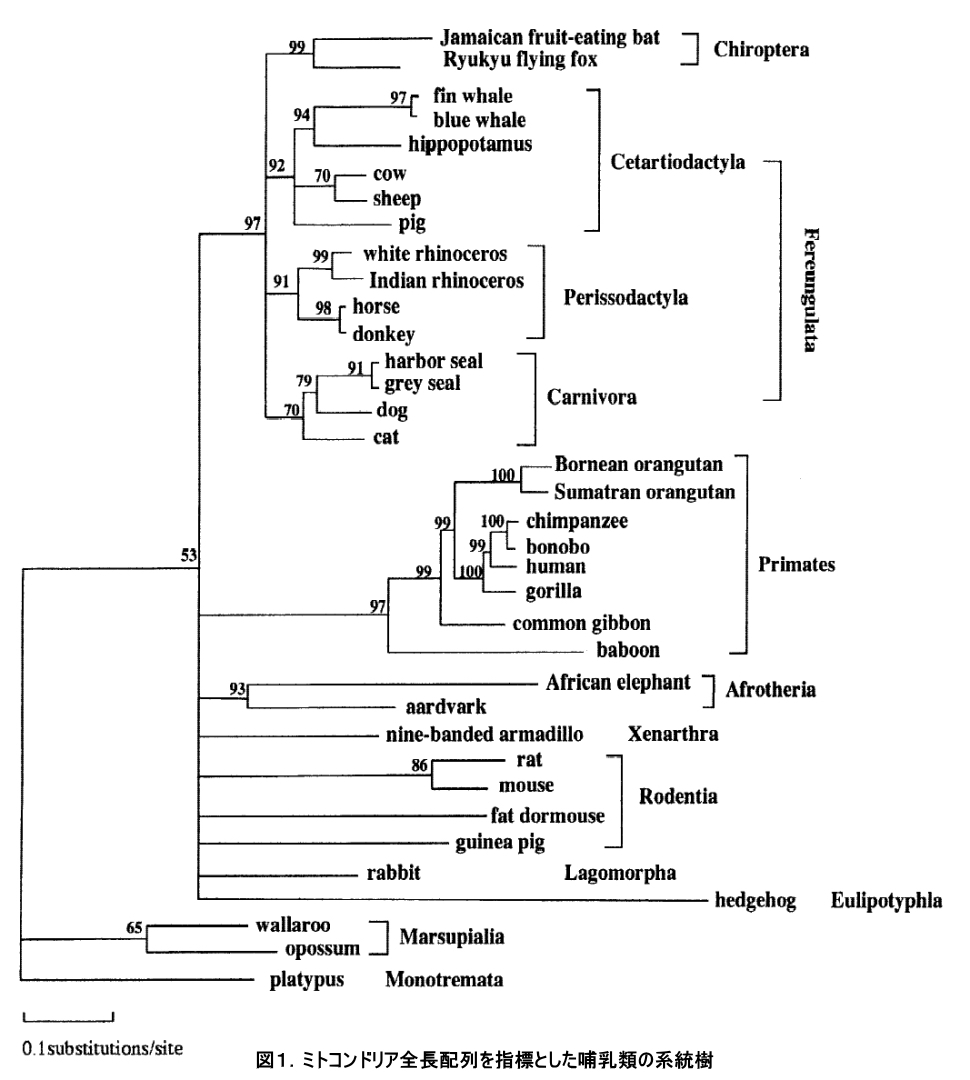
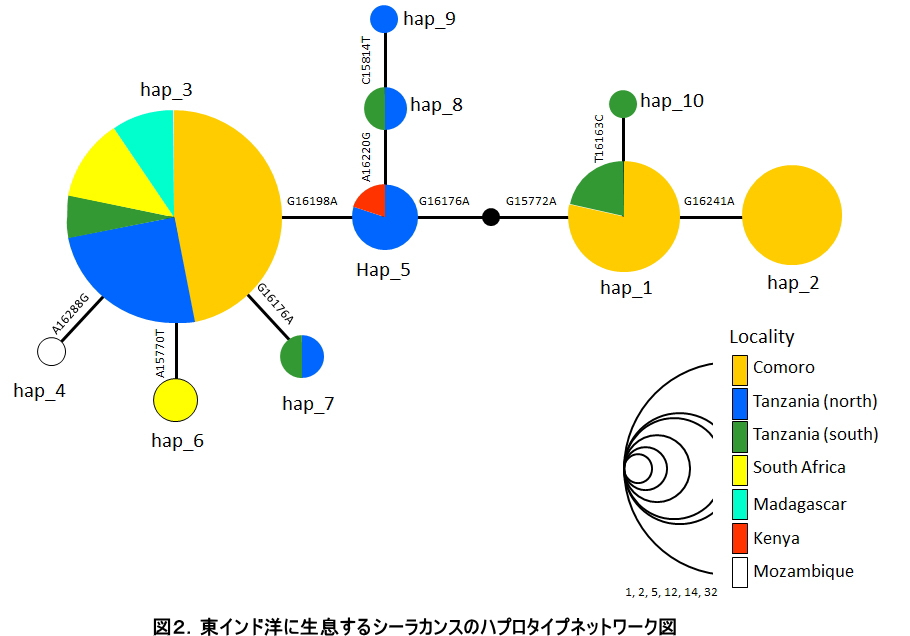
「生きた化石」と呼ばれ、研究者のみならず一般にも広く知られているシーラカンスは、1938年に南アフリカのカルムナ川河口において世界で初めて生存個体の存在が確認されました。そして、その驚くべき発見以降、多くの研究者がこのシーラカンスに注目してきましたが、その営巣地については長い間議論が続いてきました。シーラカンスを新種として記載したJLB. Smithは、2体目のシーラカンスが見つかったコモロ諸島に繁殖集団が存在すると結論付け、それ以外の場所で稀に発見されるシーラカンスについては、強い海流に流されて漂着してきた「迷子」に過ぎないと考えていました(図1参照)。つまり、コモロ諸島からアフリカ大陸に向けては強い南赤道海流が存在し、さらに大陸に沿ってモザンビーク海流が南下、東アフリカ沿岸海流が北上しているため、泳ぎの不得意なシーラカンスが稀にコモロ諸島からアフリカ大陸沿岸各地に向けて流されてしまったのではないかと考えたのです。ところが、我々が2003年から2008年にかけてタンザニア漁師によって混獲された計23個体のシーラカンスに加え、コモロ産シーラカンス2個体分のミトコンドリアゲノム全長DNA配列を決定し、それらを既に報告されているコモロ産シーラカンス38個体分のDNA配列と合わせて集団遺伝学的な手法を用いて解析したところ、タンザニア北部沿岸域に生息するシーラカンス集団が、コモロ諸島のものとは遺伝的に分化していることが統計的に有意に示されました。さらに、この2集団の分岐年代を推定したところ、もっとも少なく見積もっても 20万年前には既に集団が分かれていたことが明らかとなったのです。つまり本研究によって、タンザニア北部に生息するシーラカンスは単なるコモロ諸島からの迷子集団ではなく、独自にそこを営巣地とする繁殖集団であるとする新たな知見を示すことができたのです (Nikaido et al. 2011 PNAS) 。
シーラカンス全ゲノム計画
シーラカンスは1938年に南アフリカのカルムナ川河口において世界で初めて生存個体が確認されて以降、生物学的に重要な問題を明らかにするために数多くの研究者が注目してきました。まず、「生きた化石」としても知られている現生のシーラカンスが、数億年も前の化石種とほとんど形態が変わっていない理由については、現在もまったく明らかにされていません。またシーラカンスは、脊椎動物の進化の過程において陸上化を果たしたグループともっとも近縁である種の1つであるため、水中から陸上への段階的な進化をDNAレベルで研究するのにもっとも適した生物であると期待されてきました。ただ、シーラカンスは非常に希少な生物であるため、保存状態の良好な組織を必要とするようなDNAレベルでの研究はほとんど進められておりませんでした。その状況の中、2004年に東京工業大学の岡田典弘教授(当時)は、タンザニア沿岸において地元の漁師によって混獲された後に研究目的で冷凍保管されていたシーラカンス標本を、タンザニア水産研究所(TAFIRI)から寄贈される運びとなりました。これによってシーラカンスのDNAレベルでの大規模な共同研究が始まり、その後もワシントン条約(CITES)の取り決めにのっとり、数多くのシーラカンス標本が日本へ輸入され、全ゲノム概要配列の決定に関する一大プロジェクトが始まることになりました。
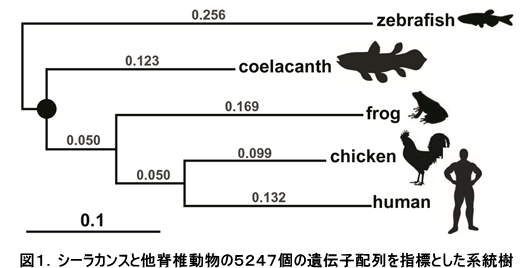
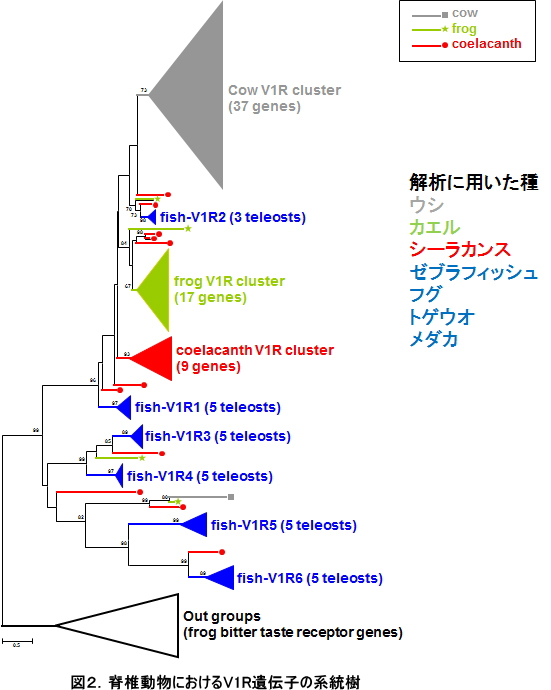
我が国におけるゲノム研究の中核をなすグループが集結してシーラカンスゲノムコンソーシアムを組織し、まず2007年にタンザニア沿岸で混獲されたシーラカンス雌個体の胎内から見つかった稚魚について、次世代シーケンサーを用いてその全ゲノム概要配列(全長約27億塩基対)を新規に決定することに成功し、続いてタンザニア産2頭とコモロ産1頭、さらにはインドネシアにおいて捕獲されたシーラカンス個体の成魚についても、全ゲノムDNA配列の決定をおこないました。この全ゲノム配列の構築には東京工業大学において独自に開発されたアッセンブラーであるPLATANUSを用いました。現存するシーラカンス2種の全ゲノム決定は世界で初めての成果になります。これらのデータを用いた大規模な比較ゲノム解析の結果、まずゲノムの進化速度が極めて遅いことが明らかとなりました(図1参照)。これまでの研究では、遺伝子のアミノ酸レベルでの進化速度が遅いことが示唆されていましたが、本研究ではDNAの進化速度そのものが遅いことを示す、世界で初めてのデータになります。これは、シーラカンスが「生きた化石」としてその形態を進化の過程においてほとんど変えていないことの原因の1つである可能性を示唆しています。また、シーラカンスの全ゲノム配列の中でも特に四肢形成や嗅覚に関連する遺伝子を魚類や四足動物(カエルや哺乳類などに代表される陸上化を遂げたグループ)と比較したところ、魚類には存在せず四足動物に特徴的であるため陸上化に深く関与していると考えられてきた遺伝子の多くが、水中に生息するシーラカンスのゲノム中にも存在することが明らかとなりました(図2参照)。四肢や嗅覚器官は、陸上化という大きな環境変化に伴ってダイナミックな進化を遂げたことが知られていますが、その前段階における祖先のゲノムにはすでに陸上化に必要な遺伝子が存在していたことが分かり、これは大規模な適応進化を可能にするDNAレベルでのメカニズムを明らかにする上で非常に重要な知見を与えるものです。さらに、本研究においては、シーラカンスの遺伝的多様性が他の野生動物と比較して極めて低いことも明らかとなり、これは希少種シーラカンスを絶滅から救うための本格的な保全活動の推進が必要であることを強く示唆しています。この成果は2013年7月にGenome Researchに発表されました(Nikaido et al. 2013 Genome Res.)。